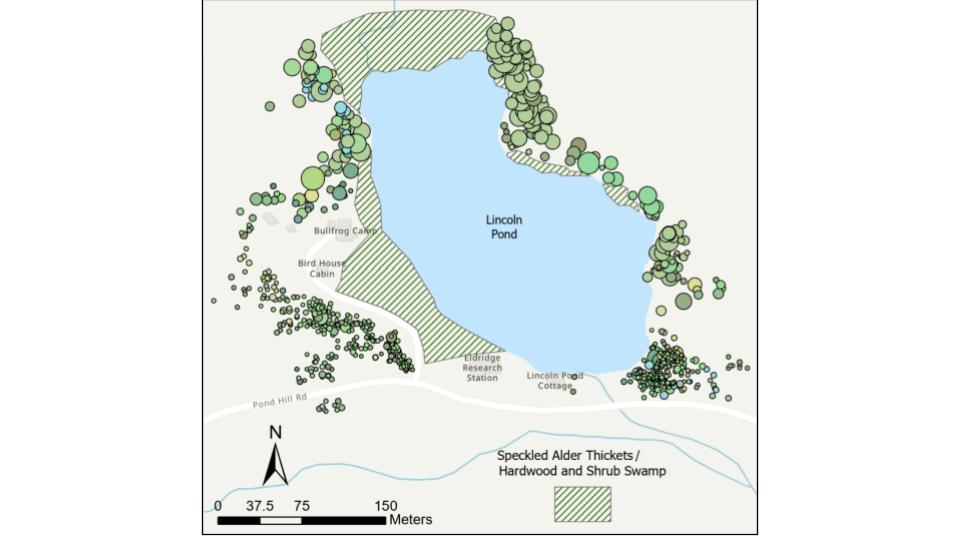
Figure A1. Map of trees fully felled (n=627) and gnawed (n=208) by beavers surrounding Lincoln Pond, summer 2024. Dot color corresponds to tree species (n=22) and dot size corresponds to tree size (Campbell & Velicer, 2024).


Ecology and Evolutionary Biology
Fall 2024, Volume 23, Issue 1
Carlina Velicer ’26, and Mark Lesser*
https://doi.org/10.47761/SQLI6534
Although beavers are considered generalist herbivores, their preference for more palatable woody plant species, such as willows and poplars, has earned them recognition as choosy, opportunistic foragers (Haarberg & Rosell, 2006; Vorel et al, 2015). A well-documented effect of selective and choosy herbivory is the proliferation of non-preferred vegetation, as herbivores with a stronger preference for native species over non-native species can encourage invasions (Augustine & McNaughton, 1998; Keane & Crawley, 2002). Previous studies have demonstrated a relationship between beaver foraging preferences and invasions of non-native trees and shrubs. In Hungary, the Eurasian beaver (Castor fiber) may expedite the shift in canopy composition toward non-native hardwood shrubs through its preference for softwood species (Juhasz et al., 2022). Beavers in Ohio may aid amur honeysuckle (Lonicera maackii) invasion by selectively felling its competition (Deardorff & Gorchov, 2020). Invasive tamarisk trees (Tamarix ramosissima) thrive where beavers are abundant in the Grand Canyon National Park (Mortenson et al., 2008).
While invasive vegetation is common in the riparian ecosystems of the northeastern United States, whether the canopy gaps created by beavers promote invasions of non-native herbaceous plants and shrubs has yet to be thoroughly investigated. In this study, we examined the influence of beavers on facilitating invasive colonization and spread at the Huyck Preserve and Biological Research Station in New York. Lincoln Pond, situated within the Preserve, has a long history of beaver activity, making it an ideal location for this study (Muller-Schwarze et. al, 1983; Tevis 1949; Tevis, 1950).
Our objectives were to determine: (1) the extent to which beaver felling influenced light availability, (2) whether canopy gaps created by beaver felling facilitated the spread of non-native herbaceous plants and woody shrubs, and (3) which invasive species took advantage of these gaps. To investigate these questions, we surveyed plots centered around beaver-felled trees and unfelled controls to characterize canopy openness, soil pH, and percent abundance of invasive species. Since beaver-felled gaps increase light availability—potentially creating conditions conducive to the spread of non-native species—we hypothesized that beaver herbivory promotes the spread of invasive herbaceous plants and shrubs at the Preserve.
Study Site
This study took place at the Huyck Preserve and Biological Research Station in east-central New York State (Fig. 1a). Beavers are well-established on Lincoln Pond, with evidence of their presence first being recorded by Eugene P. Odum in 1939 (Tevis, 1950). By 1947, two beaver lodges were observed nested along the banks of the 4 hectare-pond (Odum, 1939; Tevis, 1950). Since then, a third has been established. Over this timeframe, beavers have had a marked effect on the landscape. In a preliminary survey of beaver activity surrounding the pond, 835 trees with evidence of beaver interaction (felling or gnawing) were identified (Fig. A1, Campbell & Velicer, 2024).
The north and east sides of Lincoln Pond are dominated by old-growth eastern hemlock (Tsuga canadensis). Marshes and alder (Alnus incana) thickets cling to the north, west, and, to a lesser extent, east shorelines. The southwest shore is characterized by a red pine plantation dating back to 1928 (Tevis, 1950) that is now overgrown with mixed deciduous hardwood, namely sugar maple (Acer saccharum), American hornbeam (Carpinus caroliniana), American hophornbeam (Ostrya virginiana), and white ash (Fraxinus americana).
Invasive species are actively managed on the Preserve, through the use of a variety of techniques from manual cutting to spot-torching. Fieldwork for this study was conducted in the summer of 2024 before the annual management of invasive herbaceous plants and shrubs began around the pond area.
Field Sampling
Five-meter radial plots were surveyed around a total of 38 selected trees—19 felled and 19 unfelled control trees (Fig. 1b). Felled tree plots were selected based on the initial assessment of beaver activity around Lincoln Pond (Fig. A1, Campbell & Velicer, 2024). Only woody stems greater than 15 cm were considered, as felled trees of that size created a canopy gap large enough to alter the understory light environment. Plots were selected that avoided overlaps with a hiking trail surrounding the pond to minimize the effects of increased light availability from the path. Due to the high levels of beaver activity immediately surrounding the pond, randomly selected unfelled controls were on average farther from the pond than felled trees, but still within the riparian zone.
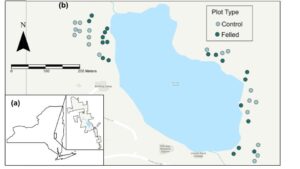 Figure 1. (a) Location of the Huyck Preserve and Biological Research Station in east-central New York State. (b) Location of surveyed plots (n=38) around Lincoln Pond, summer 2024 (plot symbols not to scale).
Figure 1. (a) Location of the Huyck Preserve and Biological Research Station in east-central New York State. (b) Location of surveyed plots (n=38) around Lincoln Pond, summer 2024 (plot symbols not to scale).
Within each plot, we estimated the percent abundance of all invasive terrestrial herbaceous plants and shrubs. Invasive trees were not present at the study location, and therefore not included in the study design. While observed in some plots, aquatic invasive species were not considered in this analysis. The abundance and growth form of some non-native species, particularly the Asiatic bittersweet (Celastrus orbiculatus), rendered counts impractical. Tree diameters were measured 20 cm from the ground, which is below beaver felling height (Belovsky, 1984; Janiszewski, 2017). We used Avenza Maps version 5.3.3 (243.1) to record tree location and a spherical densiometer to measure canopy openness. Soil samples were taken at the base of each tree, and their pH was measured with a HANNA H198107 pH meter in the lab. This controlled differences in soil pH between plot types that may have accounted for observed differences in invasive species abundance.
Statistical Analysis
Analysis of Variance (ANOVA) was performed in R version 4.3.2 (R Core Team, 2023) to test whether the mean percent abundance of invasive plants varied according to soil pH or canopy openness. Student’s t-tests were conducted to determine whether the total percent abundance of invasive species, soil pH, and canopy openness were significantly different between felled and control plots. A simple linear regression model was used to assess how well canopy openness predicted the total percent abundance of invasive species plants across all plots.
Plots centered around a total of eight different tree species were surveyed (Table 1). Two felled trees could not be identified due to significant rotting. The vast majority of felled plots were centered around deciduous trees, with eastern hemlock encompassing only ~11% of felled trees surveyed. In contrast, ~68% of control trees were eastern hemlock (Table 1). This difference can be attributed to diet preferences, as beavers generally favor more easily digestible deciduous trees over conifers (Fryxell & Doucet, 1993; Gallant et al., 2004). The study site’s high levels of beaver activity, undoubtedly a consequence of prolonged beaver residency at Lincoln Pond, rendered very few large deciduous trees left standing within the riparian zone that could serve as controls.
 Table 1. Average relative abundance and size of tree species found in the felled (n=19) and control plots (n=19).
Table 1. Average relative abundance and size of tree species found in the felled (n=19) and control plots (n=19).
There were no significant interactions between plot type, soil pH, and canopy cover, but the main effects of canopy cover and plot type were significant (Table 2, Fig. A2). However, canopy cover was highly correlated with plot type. Plots with beaver-felled trees exhibited gaps in the forest canopy, resulting in significantly higher light levels than unfelled control plots (~74% versus ~91%, respectively; Fig. 2). The total percent abundance of invasive species decreased as canopy cover increased (Fig. 3), with canopy cover explaining ~41% of the variability in the total percent abundance of invasive species (p < 0.001; Table 3).
 Table 2. Percent abundance of invasive plants as a function of soil pH and canopy cover, tested using an ANOVA.
Table 2. Percent abundance of invasive plants as a function of soil pH and canopy cover, tested using an ANOVA.
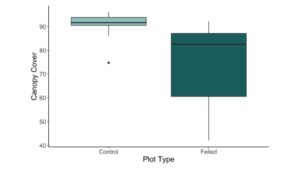 Figure 2. A boxplot of canopy cover (%) between control and felled plots (p < 0.001). Thick black line indicates the median value. Boxes represent the 1st to 3rd quartile range. Whiskers extend to the most extreme values within 1.5 times the interquartile range. Outliers are represented by black dots.
Figure 2. A boxplot of canopy cover (%) between control and felled plots (p < 0.001). Thick black line indicates the median value. Boxes represent the 1st to 3rd quartile range. Whiskers extend to the most extreme values within 1.5 times the interquartile range. Outliers are represented by black dots.
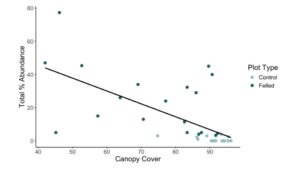 Figure 3. A scatterplot of canopy closure as a predictor of total percent abundance of invasive species plants across all plots. The line represents the fitted values from the linear regression.
Figure 3. A scatterplot of canopy closure as a predictor of total percent abundance of invasive species plants across all plots. The line represents the fitted values from the linear regression.
 Table 3. Estimated regression parameters, standard errors, and p-values for the simple linear regression model (R Square = 0.4148).
Table 3. Estimated regression parameters, standard errors, and p-values for the simple linear regression model (R Square = 0.4148).
The total percent abundance of invasive herbaceous plants and shrubs was significantly higher and more variable in beaver-felled plots than in controls (p < 0.001; Fig. 4, Table 4). However, of the ten invasive species identified, only Asiatic bittersweet, honeysuckles (Lonicera spp.), and Japanese barberry were significantly more prevalent in felled plots than in control plots (Figure 5; Table 5). Thus, only a small portion of the invasive species surveyed are drivers of the difference in invasive species prevalence by plot type.
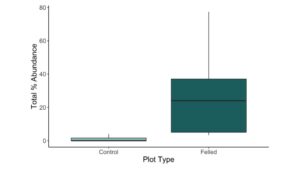 Figure 4. A boxplot of total percent abundance of invasive herbaceous plants and shrubs between control and felled plots (p < 0.001). The thick black line indicates the median value. Boxes represent the 1st to 3rd quartile range. Whiskers extend to the most extreme values within 1.5 times the interquartile range. Outliers are represented by black dots.
Figure 4. A boxplot of total percent abundance of invasive herbaceous plants and shrubs between control and felled plots (p < 0.001). The thick black line indicates the median value. Boxes represent the 1st to 3rd quartile range. Whiskers extend to the most extreme values within 1.5 times the interquartile range. Outliers are represented by black dots.
 Table 4. T-test for differences in total percent abundance of invasive species, canopy openness, and soil pH between felled and control plots.
Table 4. T-test for differences in total percent abundance of invasive species, canopy openness, and soil pH between felled and control plots.
 Figure 5. A boxplot of percent abundance of observed invasive herbaceous plant and shrub species between control and felled plots (p < 0.001). A thick black line indicates the median value. Boxes represent the 1st to 3rd quartile range. Whiskers extend to the most extreme values within 1.5 times the interquartile range. Outliers are represented by black dots.
Figure 5. A boxplot of percent abundance of observed invasive herbaceous plant and shrub species between control and felled plots (p < 0.001). A thick black line indicates the median value. Boxes represent the 1st to 3rd quartile range. Whiskers extend to the most extreme values within 1.5 times the interquartile range. Outliers are represented by black dots.
 Table 5. Student’s t-tests comparing the mean percent abundance of each invasive herbaceous plant and shrub species in felled versus control plots.
Table 5. Student’s t-tests comparing the mean percent abundance of each invasive herbaceous plant and shrub species in felled versus control plots.
Beaver-created canopy gaps exhibited significantly increased understory light availability, a finding consistent with previous observations in the midwestern United States (Barnes & Dibble, 1988; Johnston & Naiman, 1990). As predicted, higher light levels, resulting from beaver felling, were associated with a greater prevalence of invasive herbaceous plants and woody shrubs. This observed correlation between canopy openness and invasive species abundance reinforces light as a crucial resource for invasive species proliferation (Blair et al., 2010). Other studies have also highlighted the association between increased light availability due to canopy disturbance and the spread of invasive plants (Belote et al., 2008; Eschtruth & Battles, 2009; Lee & Thompson, 2012).
The dominance of Asiatic bittersweet, honeysuckles, and Japanese barberry in driving the observed differences in total invasive species abundance between plot types suggests that not all invasive plants are equally skilled in exploiting the gaps created by beavers. The success of the most prevalent invader in felled plots, Asiatic bittersweet, is likely bolstered by its rapid seed production, high germination rates, and rapid growth in high-light environments (Ellsworth, 2003). The hardy invader may endure years in heavily shaded understory before rapidly colonizing canopy gaps created by disturbance—likely giving it an advantage over other invaders such as common St. Johnswort (Hypericum perforatum), whose seedlings are small and slow-growing, or even common buckthorn (Rhamnus cathartica), whose growth rate is considered medium to fast (Campbell, 1985; Dirr, 1998; Patterson, 1974).
The honeysuckles found on the Preserve are also fast-growing and tolerant of a wide range of light conditions. Their seeds are readily dispersed by white-tailed deer populations, and, like the seeds of Asiatic bittersweet and Japanese barberry, by frugivorous birds. Morrow’s honeysuckle (Lonicera morrowii), one of the Preserve’s invasive honeysuckle species, can germinate beneath 2 inches of leaf litter (Hidayati, 2000). This ability may provide it the edge needed to invade dense hemlock forest, where thick leaf litter typically inhibits colonization by most herbaceous plants and shrubs. While not a particularly fast grower, Japanese barberry has been observed exploiting canopy gaps in hemlock forest, and its seedling survival and growth rate is also positively correlated with light level (D’Appollonio, 2006; Eschtruth & Battles, 2009; Silander, 1999).
In contrast to the more successful invaders, garlic mustard is most competitive in well-shaded understory (Dhillion, 1999), and was only observed in a single control plot in this study. Reed canary grass (Phalaris arundinacea) and timothy grass (Phleum pratense) were restricted to plots on a thin strip of wetland sandwiched between the pond’s edge and the beginning of hemlock forest. Thus, the non-significant increase in the percent abundance of these grasses in felled plots may be influenced by the slight difference in distance from the pond between felled and control plots.
The lack of significant interactions between soil pH and invasive species abundance further underscores the dominant role of light availability in facilitating non-native plant invasions. Soil pH was measured to account for any possible confounding effects of soil chemistry on invasive colonization across plot types. Although, it is possible that the increased presence of invasive plants may also be a contributing factor to the higher soil pH levels observed in felled plots. Asiatic bittersweet and Morrow’s honeysuckle have been observed to significantly increase soil pH (Hicks, 2004).
While beavers are typically recognized as wetland engineers with positive impacts on biodiversity and are often introduced to habitats as part of ecological restoration efforts (Law et al., 2019; Smith & Mather, 2013; Stringer & Gaywood, 2016), this research sheds light on an understudied aspect of their ecological impact: their potential to aid terrestrial plant invasions. Common invaders in the northeastern United States, such as Asiatic bittersweet and Japanese barberry, are known to monopolize resources and reduce biodiversity by out-competing native vegetation (Delisle & Parshall, 2018; Dibble & Rees, 2005; Ellsworth, 2003; McNab and Meeker, 1987).
The findings of this study may be context-dependent based on forest type and the legacy of invasive plants in the area. More studies, particularly across larger sites without annual invasive plant management, are needed to gain a more comprehensive understanding of the possible long-term impacts of beaver felling on plant invasions and implications for invasive plant management at a broader scale. Investigating the relationship between near-shore tree felling and aquatic invasive species abundance also warrants further consideration. Beavers’ influence on invasive plant spread may be important to consider in wetland restoration and management.
Augustine, D. J., & McNaughton, S. J. (1998). Ungulate Effects on the Functional Species Composition of Plant Communities: Herbivore Selectivity and Plant Tolerance. The Journal of Wildlife Management, 62(4), 1165–1183. https://doi.org/10.2307/3801981
Barnes, W. J., & Dibble, E. (1988). The effects of Beaver in Riverbank Forest Succession. Canadian Journal of Botany, 66(1), 40–44. https://doi.org/10.1139/b88-005
Belote, R. T., Jones, R. H., Hood, S. M., & Wender, B. W. (2008). Diversity–invasibility across an experimental disturbance gradient in Appalachian forests. Ecology, 89(1), 183–192. https://doi.org/10.1890/07-0270.1
Belovsky, G. E. (1984). Summer Diet Optimization by Beaver. The American Midland Naturalist, 111(2), 209–222. https://doi.org/10.2307/2425316
Blair, B. C., Letourneau, D. K., Bothwell, S. G., & Hayes, G. F. (2010). Disturbance, resources, and exotic plant invasion: Gap Size Effects in a redwood forest. Madroño, 57(1), 11–19. https://doi.org/10.3120/0024-9637-57.1.11
Campbell, M. H. (1985). Germination, emergence and seedling growth of Hypericum perforatum L. Weed Research, 25(4), 259-266. https://doi.org/10.1111/j.1365-3180.1985.tb00643.x
Campbell, R., Velicer, C. (2024, August 8). Just How Busy are the Beavers? An Overview of Beaver Activity Around Lincoln Pond. [Presentation] Thursday Night Lecture Series, Huyck Preserve and Biological Research Station, Rensselaerville, NY, United States.
D’Appollonio, J. (2006). Regeneration strategies of Japanese barberry (Berberis thunbergii DC.) in coastal forests of Maine. https://digitalcommons.library.umaine.edu/etd/433
Deardorff, J. L., & Gorchov, D. L. (2020). Beavers cut, but do not prefer, an invasive shrub, Amur honeysuckle (Lonicera maackii). Biological Invasions, 23(1), 193–204. https://doi.org/10.1007/s10530-020-02365-8
Dibble, A. C., & Rees, C. A. (2005). Does the lack of reference ecosystems limit our science? A case study in nonnnative invasive plants as forest fuels. Journal of Forestry, 103(7), 329–338. https://doi.org/10.1093/jof/103.7.329
Delisle, Z. J., & Parshall, T. (2018). The Effects of Oriental Bittersweet on Native Trees in a New England Floodplain. Northeastern Naturalist, 25(2), 188–196. https://www.jstor.org/stable/26577814
Dhillion, S. S., & Anderson, R. C. (1999). Growth and Photosynthetic Response of First-Year Garlic Mustard (Alliaria petiolata) to Varied Irradiance. The Journal of the Torrey Botanical Society, 126(1), 9–14. https://doi.org/10.2307/2997250
Dirr, M. (1998). Manual of woody landscape plants: Their identification ornamen- tal characteristics, culture, propagation and uses (5th ed.). Stipes Publishing.
Donkor, N. T., & Fryxell, J. M. (1999). Impact of beaver foraging on structure of lowland boreal forests of Algonquin Provincial Park, Ontario. Forest Ecology and Management, 118(1–3), 83–92. https://doi.org/10.1016/s0378-1127(98)00487-3
Ellsworth, J. W. (2003). Controls on the Establishment and Early Growth of Oriental Bittersweet (Celastrus Orbiculatus Thunb.), an Invasive Woody Vine (Doctoral dissertation, University of Massachusetts at Amherst).
Eschtruth, A. K., & Battles, J. J. (2009). Assessing the relative importance of disturbance, herbivory, diversity, and propagule pressure in exotic plant invasion. Ecological Monographs, 79(2), 265–280. https://doi.org/10.1890/08-0221.1
Fryxell, J. M., & Doucet, C. M. (1993). Diet Choice and the functional response of Beavers. Ecology, 74(5), 1297–1306. https://doi.org/10.2307/1940060
Gallant, D., Bérubé, C. H., Tremblay, E., & Vasseur, L. (2004). An extensive study of the foraging ecology of beavers (Castor canadensis) in relation to habitat quality. Canadian Journal of Zoology, 82(6), 922–933. https://doi.org/10.1139/z04-067
Haarberg, O., & Rosell, F. (2006). Selective foraging on woody plant species by the Eurasian beaver (Castor fiber) in Telemark, Norway. Journal of Zoology, 270(2), 201–208. https://doi.org/10.1111/j.1469-7998.2006.00142.x
Hicks, S. L. (2004). The effects of invasive species on soil biogeochemistry. Science Daily. Hampshire College, Amherst, MA, 1002.
Hidayati, S. N., Baskin, J. M., & Baskin, C. C. (2000). Dormancy-breaking and germination requirements of seeds of four Lonicera species (Caprifoliaceae) with underdeveloped spatulate embryos. Seed Science Research, 10(4), 459-469.
Janiszewski, P., Kolasa, S., & Strychalski, J. (2017). The preferences of the European beaver Castor fiber for trees and shrubs in riparian zones. Applied Ecology & Environmental Research, 15(4).
Johnston, C. A., & Naiman, R. J. (1990). Browse selection by Beaver: Effects on riparian forest composition. Canadian Journal of Forest Research, 20(7), 1036–1043. https://doi.org/10.1139/x90-138
Juhász, E., Bede‐Fazekas, Á., Katona, K., Molnár, Z., & Biró, M. (2022). Foraging decisions with conservation consequences: Interaction between beavers and invasive tree species. Ecology and Evolution, 12(5). https://doi.org/10.1002/ece3.8899
Keane, R. M., & Crawley, M. J. (2002). Exotic plant invasions and the enemy release hypothesis. Trends in Ecology & Evolution, 17(4), 164–170. https://doi.org/10.1016/s0169-5347(02)02499-0
Law, A., Levanoni, O., Foster, G., Ecke, F., & Willby, N. J. (2019). Are beavers a solution to the freshwater biodiversity crisis? Diversity and Distributions, 25(11), 1763–1772. https://doi.org/10.1111/ddi.12978
Lee, T. D., & Thompson, J. H. (2012). Effects of logging history on invasion of eastern white pine forests by exotic glossy buckthorn (Frangula Alnus P. Mill.). Forest Ecology and Management, 265, 201–210. https://doi.org/10.1016/j.foreco.2011.10.035
Loh, R. K., & Daehler, C. C. (2008). Influence of woody invader control methods and seed availability on native and invasive species establishment in a Hawaiian forest. Biological Invasions, 10(6), 805–819. https://doi.org/10.1007/s10530-008-9237-y
McNab, W. H., & Meeker, M. (1987). Oriental bittersweet: A growing threat to hardwood silviculture in the Appalachians. Northern Journal of Applied Forestry, 4(4), 174–177. https://doi.org/10.1093/njaf/4.4.174
Mortenson, S. G., Weisberg, P. J., & Ralston, B. E. (2008). Do beavers promote the invasion of non-native Tamarix in the Grand Canyon Riparian Zone? Wetlands, 28(3), 666–675. https://doi.org/10.1672/07-142.1
Muller-Schwarze, D., S. Heckman and B. Stagge. 1983. Behavior of free-ranging beaver (Castor canadensis) at scent marks. Acta Zool. Fennica 174:111-113.
Odum, E. P. (1943). The Vegetation of the Edmund Niles Huyck Preserve, New York. The American Midland Naturalist, 29(1), 72–88. https://doi.org/10.2307/2420980
Patterson, D. T. (1974). The ecology of oriental bittersweet, Celastrus orbiculatus, a weedy introduced ornamental vine. Duke University.
R Core Team (2023). R (4.3.2): A Language and Environment for Statistical Computing. Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/.
Rosell, F., Bozsér, O., Collen, P., & Parker, H. (2005). Ecological impact of beavers castor fiber and castor canadensis and their ability to modify ecosystems. Mammal Review, 35(3–4), 248–276. https://doi.org/10.1111/j.1365-2907.2005.00067.x
Silander, J. A., & Klepeis, D. M. (1999). The invasion ecology of Japanese barberry (Berberis thunbergii) in the New England landscape. Biological Invasions 1, 189–201 (1999). https://doi.org/10.1023/A:1010024202294
Smith, J. M., & Mather, M. E. (2013). Beaver dams maintain fish biodiversity by increasing habitat heterogeneity throughout a low‐gradient Stream Network. Freshwater Biology, 58(7), 1523–1538. https://doi.org/10.1111/fwb.12153
Stringer, A. P., & Gaywood, M. J. (2016). The impacts of beavers Castor spp. on biodiversity and the ecological basis for their reintroduction to Scotland, UK. Mammal Review, 46(4), 270–283. https://doi.org/10.1111/mam.12068
Tevis, L. 1949. A scientist watches the beaver. Bulletin to the Schools (State University of New York) 35:225-229.
Tevis, L. (1950). Summer Behavior of a Family of Beavers in New York State. Journal of Mammalogy, 31(1), 40–65. https://doi.org/10.2307/1375473
Vorel, A., Válková, L., Hamšíková, L., Maloň, J., & Korbelová, J. (2015). Beaver foraging behaviour: Seasonal foraging specialization by a choosy generalist herbivore. Behavioral Ecology and Sociobiology, 69(7), 1221–1235. https://doi.org/10.1007/s00265-015-1936-7
Figure A1. Map of trees fully felled (n=627) and gnawed (n=208) by beavers surrounding Lincoln Pond, summer 2024. Dot color corresponds to tree species (n=22) and dot size corresponds to tree size (Campbell & Velicer, 2024).
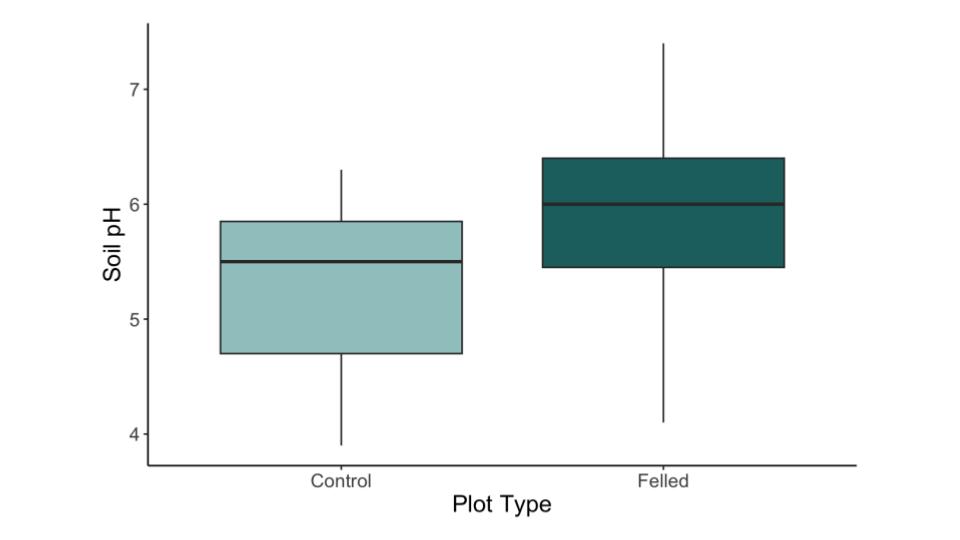
Figure A2. Boxplot of soil pH between control and felled plots (p < 0.05). Thick black line indicates the median value. Boxes represent the 1st to 3rd quartile range. Whiskers extend to the most extreme values within 1.5 times the interquartile range. Outliers represented by black dots.
I am a third-year undergraduate majoring in ecology and evolutionary biology and minoring in environmental humanities. I conducted this research during my time as an Odum field ecology intern at the Huyck Preserve and Biological Research Station, located near Albany. My interests lie in research that informs conservation efforts and our understanding of ecological responses to abiotic and biotic disturbances such as climate change, habitat loss, and invasions of non-native species.
Velicer, C. and Lesser, M. (2024). Engineering invasion: Do canopy gaps created by the North American beaver (Castor canadensis) facilitate terrestrial plant invasions? University of Rochester, Journal of Undergraduate Research, 23(1). https://doi.org/10.47761/SQLI6534
JUR | Creative Commons Attribution 4.0 BY International License